
Revisión académica de investigación universitaria
Dr. Alfonso Barbosa Domínguez
Magister en Fisiología del deporte
PHD en Neurofisiología
Post-PHD en Epigenética
Contexto
Recientemente descubrimos que la exposición prolongada al óxido nítrico (NO) desencadena la biogénesis mitocondrial en células y tejidos de mamíferos mediante la activación del guanilato ciclasa y la generación de cGMP. En este trabajo, informamos que la biogénesis mitocondrial dependiente de NO/cGMP se asocia con una respiración acoplada mejorada y un mayor contenido de ATP en células U937, L6 y PC12. El aumento observado en el contenido de ATP dependió completamente de la fosforilación oxidativa, ya que la formación de ATP por glucólisis no varió. El cerebro, el riñón, el hígado, el corazón y el músculo gastrocnemio de ratones mutantes nulos de la NO sintasa endotelial mostraron un contenido mitocondrial notablemente reducido, asociado con un consumo de oxígeno y un contenido de ATP significativamente menores. En estos tejidos, los análisis ultraestructurales revelaron mitocondrias significativamente más pequeñas. Además, se observó una reducción significativa en el número de mitocondrias en la región subsarcolemal del músculo gastrocnemio. Concluimos que NO/cGMP estimula la biogénesis mitocondrial, tanto in vitro como in vivo, y que esta estimulación está asociada con una mayor función mitocondrial, lo que resulta en una mayor formación de ATP.
El óxido nítrico (NO) es una molécula de señalización ubicua involucrada en varias funciones fisiológicas. También puede mediar efectos deletéreos que se vuelven evidentes después de su formación inapropiada o excesiva. Algunos de los efectos fisiológicos y patológicos del NO resultan de sus acciones a nivel mitocondrial. En concentraciones nano molares, el NO se une al citocromo c (Cyt c) oxidasa (complejo IV, COX-IV), la enzima terminal en la cadena de transporte de electrones mitocondrial, inhibiendo su actividad de manera reversible y en competencia con O2 (3–5 ) . La regulación dependiente de NO de la respiración mitocondrial y el potencial de membrana contribuye a la detección aguda de O2 por las células. Además, la unión del NO a la COX-IV induce un cambio a la glucólisis en las células competentes, la redistribución del O₂ y la regulación de los niveles del factor inducible por hipoxia 1α, contribuyendo así a la adaptación a largo plazo a condiciones hipóxicas. Por el contrario, altas concentraciones de NO inhiben persistentemente los complejos I y II de la cadena respiratoria, así como las enzimas de la vía glucolítica y el ciclo de Krebs, lo que provoca desequilibrio metabólico y daño celular.
Implicaciones en el deporte y fisiológica:
Mecanismos moleculares:
- Serina y PHGDH: Un estudio de 2024 en Science Advances revela que la deficiencia de (fosfoglicerato deshidrogenasa) potencia la biogénesis mitocondrial en macrófagos inflamatorios, sugiriendo que la síntesis de serina controla este proceso vía mtROS y HIF-1α.
- TAZ: Se ha identificado a TAZ (coactivador transcripcional con motivo de unión a PDZ) como un nuevo estimulador de la biogénesis mitocondrial en músculo esquelético, vinculado al ejercicio, a través de la vía Rheb/mTOR.
- NO/cGMP: Investigaciones demuestran que el óxido nítrico (NO) y el cGMP estimulan la biogénesis mitocondrial, mejorando la respiración y el ATP, con implicaciones en varios tejidos.
Implicaciones en enfermedades y salud:
- Neuro protección y neurodegeneración: La biogénesis mitocondrial mejorada promueve la neuro protección (células ganglionares de retina). También se ha demostrado que la biogénesis mediada por PGC-1α es crucial para la recuperación neuronal tras daño.
- Cáncer: La biogénesis mitocondrial está implicada en la progresión del cáncer, con el factor TFAM y PGC-1β jugando roles complejos en tumores gastrointestinales.
- Obesidad y ejercicio: El ejercicio moderado previene la pérdida mitocondrial en la obesidad, de forma independiente a la vía AMPK/PGC-1α, implicando a la proteína de fisión DRP-1 para mejorar el control de calidad mitocondrial.
- Enfermedades cardiovasculares: Las alteraciones en la biogénesis mitocondrial y la energía de los cardiomiocitos se vinculan a enfermedades cardiovasculares, siendo PGC-1α y eNOS dianas terapéuticas.
Reguladores clave: Marcadores Biológicos y genéticos.
- PGC-1α/β, NRF1/2, TFAM: Estos factores siguen siendo centrales en la regulación de la biogénesis mitocondrial, aunque nuevos estudios están desvelando vías alternativas y complejas (como la no dependencia de AMPK/PGC-1α en el ejercicio
Recientemente hemos demostrado que la exposición a largo plazo de células en cultivo a bajas concentraciones de NO induce la biogénesis mitocondrial. Este proceso está mediado por cGMP, resultante de la activación dependiente de NO del guanilato ciclasa “soluble” (sGC), e implica una mayor expresión del coactivador 1α del receptor activado por el proliferador de peroxisomas γ (PGC-1α), el factor respiratorio nuclear 1 (NRF-1) y el factor de transcripción mitocondrial A (Tfam). Otros estímulos biogenéticos mitocondriales también aumentan la expresión de estos factores de transcripción. La biogénesis mitocondrial dependiente de NO es importante para el metabolismo celular y tisular, como lo demuestran los estudios en ratones deficientes en la sintasa de NO endotelial (eNOS). La biogénesis mitocondrial reducida en los tejidos de estos animales se asocia con un menor gasto energético y un mayor peso corporal. En este estudio, hemos caracterizado el estado funcional de las mitocondrias generadas por la acción del NO y hemos demostrado que la biogénesis mitocondrial dependiente de NO/cGMP produce mitocondrias funcionalmente activas, en términos de función respiratoria y actividad metabólica, en varias células de mamíferos, así como en tejidos animales.
Materiales y métodos
Materiales. Los siguientes reactivos se adquirieron según lo indicado: (Z)-1-[2-(2-aminoetil)- N -(2-amonioetil) amino]diazen-1-io-1,2 diolato (DETA-NO) y H -(1,2,4)-oxadiazolo [4,3-] quinoxalin-1-ona (ODQ) de Alexis Italia (Florencia, Italia); BAY 41-2272 de Bayer (Wuppertal, Alemania); mAb primarios antisubunidad IV de COX-IV de Molecular Probes; mAb anti-Cyt c de Pharmingen; e IgG de cabra anticonejo marcada con isotiocianato de fluoresceína de The Jackson Laboratory. El clon fetal III se obtuvo de HyClone–Celbio (Milán). Todos los demás reactivos de cultivo celular se obtuvieron de GIBCO (Basilea). El cianuro de carbonilo p -trifluorometoxifenilhidrazona (FCCP) y todos los demás reactivos se obtuvieron de Sigma-Aldrich (Milán).
Animales y tejidos. Ratones machos de 8 a 10 semanas de edad WT y mutantes nulos de eNOS (eNOS –/–) ( 16 ) se alojaron en la Instalación Libre de Patógenos en el Departamento de Ciencias Preclínicas de la Universidad de Milán. Los animales fueron tratados de acuerdo con las directrices de la Comunidad Europea y con la aprobación del Comité Ético Institucional. El día de los experimentos, los animales fueron sacrificados por dislocación cervical y los tejidos se aislaron inmediatamente. Se congelaron en nitrógeno líquido (para estudios de ADNmt, proteína mitocondrial y ATP), o se liberaron de tejido conectivo, grasa y vasculatura grande; se cortaron en rebanadas de 20 a 30 mg; y se mantuvo en una solución de Krebs que contenía 118 mM de NaCl, 4,7 mM de KCl, 1,5 mM de CaCl 2 , 25 mM de NaHCO 3 , 1,2 mM de KH 2 PO 4 , 1,1 mM de MgSO 4 y 10 mM de glucosa (pH 7,4) en la que se burbujeó continuamente 20 % de O 2 / 5 % de CO 2 / 75 % de N 2 a 37 °C (para los experimentos de consumo de O 2 ).
Cultivo celular y tratamientos. Las células U937, L6 y PC12 se cultivaron esencialmente como se describe (22–24 ) . En el día 0, las células U937 se suspendieron a una densidad de 3 × 104 células por ml, mientras que L6 y PC12 se sembraron a una densidad de 1 × 105 células por mm2. DETA-NO, 8 Br-cGMP, BAY 41-2272 y ODQ se añadieron a los cultivos en varias combinaciones, como se indica en Resultados, una vez al día durante 6 días. Después de este tiempo, para minimizar los posibles efectos de la privación de nutrientes en las células, se lavaron una vez en medio de cultivo fresco sin fármaco y se equilibraron en él durante 3 h más a 37 °C. A continuación, las células se recogieron y se lavaron dos veces en PBS. Para los experimentos de consumo de O₂ y citometría de flujo, las células se suspendieron (10⁻¹ células por ml) en un tampón con 118 mM de NaCl, 4,8 mM de KCl, 1,2 mM de KH₂PO₃, 1,2 mM de MgSO₃, 1 mM de CaCl₂, 10 mM de glucosa y 25 mM de Hepes (pH 7,2). Para el resto de los procedimientos experimentales, las células se sedimentaron por centrifugación y se mantuvieron a –80 °C hasta su uso.
Análisis de mtADN y proteínas. Las mitocondrias se aislaron de células y tejidos cultivados como se describe. La extracción y purificación de mtADN de mitocondrias lisadas se llevó a cabo como se describe. Se cargaron alícuotas de mtADN en gel de agarosa teñido con bromuro de etidio (1,2%) y se analizaron utilizando el densitómetro QuickImage (Packard). Los niveles de mtADN se normalizaron al contenido de proteína, que se midió utilizando el procedimiento de ensayo de proteína de ácido bicinconínico (Perbio, Bezons, Francia). El análisis del contenido celular de proteínas mitocondriales se llevó a cabo por citometría de flujo, como se describe. Brevemente, las suspensiones celulares se permeabilizaron durante 20 min a temperatura ambiente en presencia de 1% de BSA y 0,1% de saponina. Las muestras (1 × 106 células) se incubaron con los anticuerpos primarios apropiados durante 30 minutos a 4 °C. La expresión de estas moléculas se analizó mediante citometría de flujo después de teñirlas con los anticuerpos marcados con isotiocianato de fluoresceína apropiados, utilizando un clasificador de células activado por fluorescencia (FACStar Plus, Beckton Dickinson).
RT-PCR cuantitativa. La RT-PCR cuantitativa se llevó a cabo utilizando un sistema de detección de secuencias ABI Prism 7700 y TaqMan (Applied Biosystems), que utiliza la actividad de nucleasa 5′ de la TaqDNA polimerasa para generar un ensayo de análisis de ADN cuantitativo en tiempo real. Brevemente, se diseñaron sondas de oligonucleótidos específicos de genes con grupos fluorescentes 5′ y rodamina 3′ (quench) y se utilizaron para la fase de extensión de la PCR. La degradación y liberación del grupo fluorescente da como resultado fluorescencia a 518 nm, que se monitorea durante todo el proceso de amplificación. Se llevaron a cabo comparaciones con gliceraldehído-3-fosfato deshidrogenasa (control interno) y curvas estándar individuales en paralelo.
Medición del consumo de O₂. Analizamos muestras de células o tejidos de 1 ml a 37 °C en un recipiente hermético equipado con un electrodo de O₂ tipo Clark, Rank Brothers, Bottisham, Reino Unido) conectado a un registrador gráfico. El consumo celular de O₂ se midió como se describe (14). El electrodo de O₂ se calibró asumiendo que la concentración de O₂ en el medio de incubación a 37 °C era de 200 μM. El desacoplador FCCP se añadió directamente a la cubeta, mientras que la oligomicina se añadió 20 min antes de las mediciones del consumo de O₂. El contenido proteico en las muestras de células y tejidos se determinó mediante el ensayo de proteína con ácido bicinconínico.
Mediciones de ATP, lactato y actividades de enzimas glucolíticas. El contenido de ATP de células cultivadas y tejidos animales se determinó en extractos de ácido perclórico al 2,5% neutralizados con K2CO3, por HPLC de fase reversa, como se describe. El lactato se midió en el medio de cultivo después de la desproteinización utilizando dispositivos de filtro centrífugo Ultra-4 (corte, 10.000; Amicon) así como en células después de la extracción en ácido perclórico frío como se describe. Las actividades de las enzimas hexoquinasa (EC 2.7.1.1) y gliceraldehído fosfato deshidrogenasa (EC 1.2.1.13) se determinaron en lisados celulares obtenidos por sonicación (tres pulsos de 30 s a 100 W) como se describe.
Microscopía electrónica. El hígado y el músculo gastrocnemio se extrajeron cuidadosamente de ratones WT y eNOS –/–, se cortaron en trozos de ≈1 mm 3 y se colocaron en fijador helado (glutaraldehído al 2 % en tampón de cacodilato de sodio 0,1 M, pH 7,4) durante 5 h. A continuación, las muestras se lavaron exhaustivamente con tampón de cacodilato 0,1 M, se fijaron posteriormente durante 2 h con OsO 4 al 2 % / tampón de cacodilato 0,1 M, se deshidrataron en etanol, se tiñeron en bloque con acetato de uranilo y se incluyeron en Epon. Se recogieron secciones ultrafinas en rejillas de cobre, se tiñeron doblemente con acetato de uranilo y citrato de plomo y se examinaron con un microscopio electrónico de transmisión CM10 (Philips, Eindhoven, Países Bajos). Para los estudios morfométricos de las mitocondrias, se fotografiaron áreas de tejido seleccionadas aleatoriamente, derivadas de tres animales por grupo, con un aumento de 11.500 × y se analizaron con el software de imágenes de los Institutos Nacionales de la Salud. Los análisis estadísticos del área transversal de las mitocondrias y las densidades mitocondriales se realizaron con el software Prism 2.0.
Análisis estadístico. Los resultados se expresan como media ± EEM; n representa el número de experimentos individuales. El análisis estadístico se realizó mediante la prueba t de Student para variables no pareadas (bilaterales). Los asteriscos simples, dobles y triples en las tablas y figuras indican probabilidades estadísticas de p < 0,05, p < 0,01 y p < 0,001, respectivamente.
Resultados
Las células monocíticas humanas U937, los mioblastos L6 de rata y las células neurosecretoras PC12 de rata se trataron todos los días durante 6 días con varias combinaciones de los siguientes agentes: el donante de NO DETA-NO [50 μM; produciendo una concentración de NO de 98 ± 9,2 nM ( n = 3), medida con un electrodo de NO , el análogo de cGMP permeable a la membrana 8 Br-cGMP (3 mM), BAY 41-2272 [1 μM; un compuesto que activa sGC a través de un sitio independiente de NO, y el inhibidor de sGC ODQ (1 μM). Ninguno de los tratamientos afectó la viabilidad celular, según se evaluó diariamente mediante el ensayo de exclusión de azul tripán (datos no mostrados).
El tratamiento durante 6 días con DETA-NO mejoró la expresión de PGC-1α, NRF-1 y Tfam, aunque en diferentes grados, en los tres tipos celulares ( Fig. 1 ). De manera similar, aumentó el contenido de ADNmt, al igual que la expresión de las proteínas mitocondriales COX-IV y Cyt c ( Fig. 2 ). Los efectos de DETA-NO fueron imitados por 8 Br-cGMP o BAY 41-2272 y fueron prevenidos por ODQ (Fig. 1 y 2 ). Por lo tanto, la activación de sGC y la consiguiente generación de cGMP parecen ser necesarias y suficientes para promover la biogénesis mitocondrial dependiente de NO en tres tipos diferentes de células en cultivo.
A continuación, se investigó el impacto de la biogénesis mitocondrial dependiente de NO en la respiración celular. En vista de la dependencia de cGMP del proceso, utilizamos células tratadas durante 6 días con 8 Br-cGMP o BAY 41-2272, que son compuestos que comparten con NO un efecto en el sistema cGMP sin compartir sus efectos en COX-IV o en las vías de señalización independientes de cGMP. Estos tratamientos modificaron el contenido de proteína celular de manera diferente en las líneas celulares investigadas. Hubo aumentos netos de 25 ± 1,23% y 20 ± 1,21% en células U937 y 12,5 ± 0,65% y 5,7 ± 0,11% en células PC12 después de 8-Br cGMP y BAY 41-2272, respectivamente (n = 3), mientras que no se observaron cambios en el contenido neto de proteína en las células L6. Por lo tanto, los resultados se normalizaron al contenido de proteína para permitir una mejor comparación de los efectos sobre la biogénesis mitocondrial. El consumo de O 2 se midió en condiciones basales y después de la adición de FCCP (2 μM) u oligomicina (2,5 μM). FCCP desacopla completamente las mitocondrias y maximiza su capacidad respiratoria, lo que refleja la actividad máxima de transporte de electrones de las mitocondrias. Por el contrario, el inhibidor de la sintetasa de ATP F1/F0 oligomicina bloquea el consumo de O 2 vinculado a la fosforilación oxidativa, sin afectar el consumo de O 2 que ocurre a través de la fuga de protones. La comparación de los valores de consumo de O 2 en las tres condiciones experimentales permite una estimación de la respiración acoplada y desacoplada (. El tratamiento con 8 Br-cGMP o BAY 41-2272 aumentó las tasas de consumo de O 2 basal, estimulado por FCCP y resistente a oligomicina en los tres tipos de células ( Fig. 3 ). Los aumentos en las tasas de consumo de O₂ inducidos por cada compuesto en condiciones basales fueron similares en cada tipo celular a los medidos en presencia de FCCP y oligomicina. Este hallazgo demuestra que tanto el 8Br-cGMP como el BAY 41-2272 aumentaron la respiración acoplada, independientemente de si la biogénesis mitocondrial se asoció con cambios en el contenido proteico.
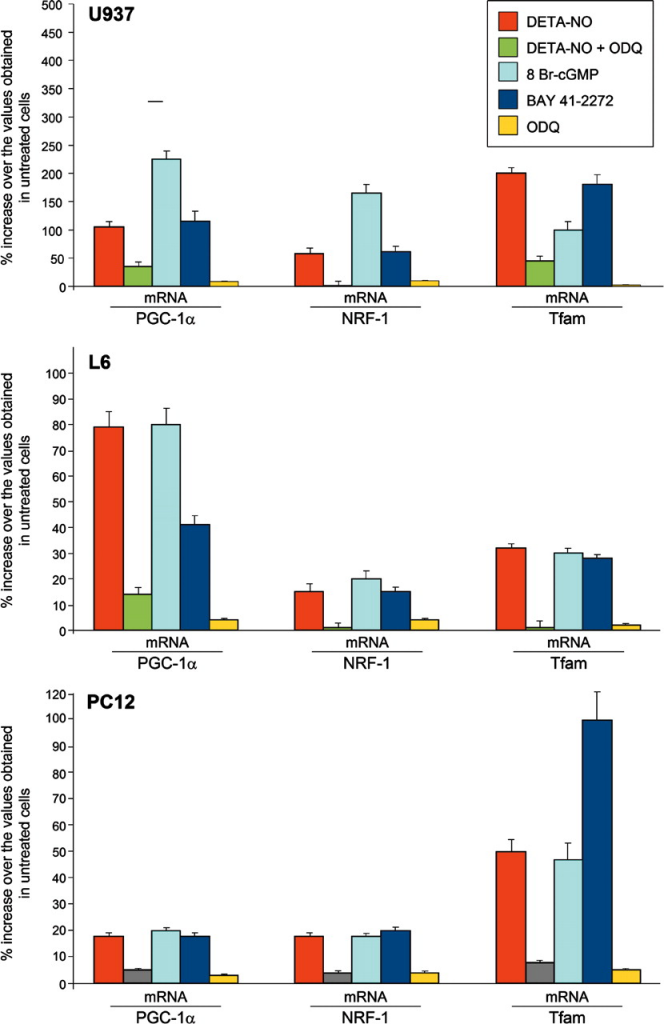
El tratamiento durante 6 días con DETA-NO mejoró la expresión de PGC-1α, NRF-1 y Tfam, aunque en diferentes grados, en los tres tipos celulares ( Fig. 1 ).
El NO desencadena la transcripción de genes implicados en la biogénesis mitocondrial en diferentes células de mamíferos mediante la generación de cGMP. Las células U937, L6 y PC12 se cultivaron durante 6 días con o sin 50 μM de DETA-NO, 3 mM de 8 Br-cGMP, 1 μM de BAY 41-2272 o 1 μM de ODQ, según se indique. Los niveles de ARNm de PGC-1α, NRF-1 y Tfam se analizaron mediante análisis de RT-PCR cuantitativa con sondas de oligonucleótidos específicos del gen. La fluorescencia específica se monitorizó durante todo el proceso de amplificación y se comparó con la fluorescencia de la gliceraldehído-3-fosfato deshidrogenasa (control interno). Los niveles de ARNm se expresaron como porcentajes de aumento sobre los valores obtenidos en células no tratadas (n = 5). Con todos los tratamientos, excepto aquellos con DETA-NO más ODQ o ODQ solo, los porcentajes de aumento sobre los valores basales fueron estadísticamente significativos (P < 0,01).
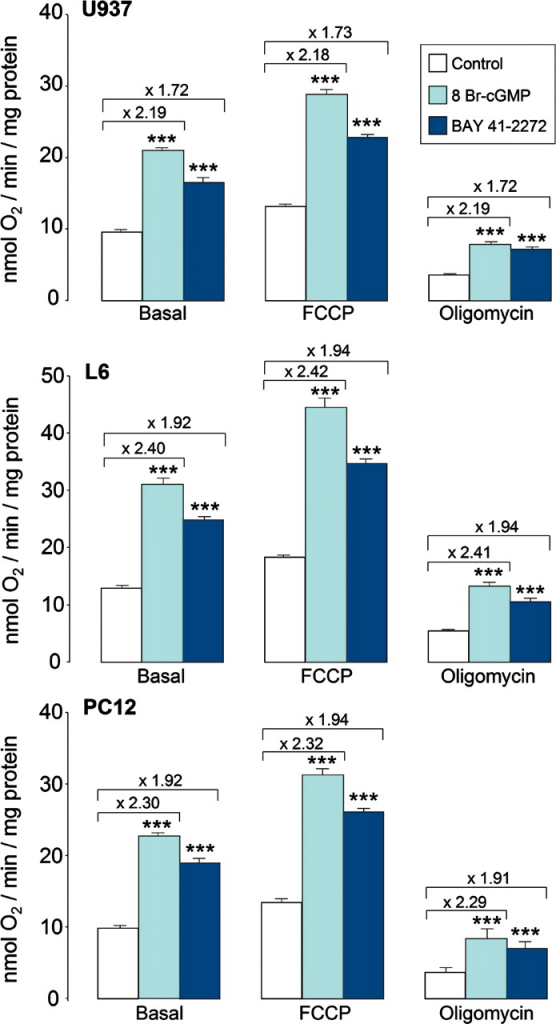
La biogénesis mitocondrial dependiente de cGMP conduce a un aumento en la respiración acoplada a la fosforilación oxidativa. Las células U937, L6 y PC12 se cultivaron durante 6 días con o sin 3 mM de 8 Br-cGMP o 1 μM de BAY 41-2272, como se indica. El consumo de O2 se midió en una cámara hermética utilizando un electrodo de O2 (como se describe en Materiales y métodos en las muestras; 107 células por muestra) de las diversas preparaciones celulares en presencia o ausencia del desacoplador mitocondrial FCCP (2 μM) o el inhibidor de la sintetasa de ATP F1/F0 oligomicina (2,5 μM). Los valores de consumo de O2 se normalizaron al contenido proteico de las células (n = 6). En cada imagen, los números sobre las barras indican el aumento en la respiración sobre el valor basal inducido por el tratamiento celular con 8 Br-cGMP o BAY 41-2272. Los asteriscos indican significancia estadística, como se describe en Materiales y métodos, frente a los controles.
A continuación, investigamos si el aumento de la respiración acoplada se asociaba a un aumento de la síntesis de ATP, y descubrimos que el tratamiento con 8 Br-cGMP o BAY 41-2272 aumentaba significativamente los niveles de ATP celular en los tres tipos celulares ( Fig. 4 A ). Los niveles de ATP dependen no solo del contenido mitocondrial, sino también de la disponibilidad de ADP y de los cambios en la demanda energética. Cabe destacar que estos parámetros se modifican significativamente durante la división celular. Aunque 8 Br-cGMP y BAY 41-2272 no modificaron la proliferación de células U937, inhibieron la proliferación de células L6 en un 34,0 ± 2,11 % y un 18,1 ± 0,83 %, y de PC12 en un 92,5 ± 5,41 % y un 80,5 ± 4,31 %, respectivamente (n = 3). La observación de que los niveles de ATP aumentaron independientemente de los efectos sobre la proliferación celular descarta la posibilidad de que el ATP se acumule debido a la menor utilización debida a la proliferación reducida.
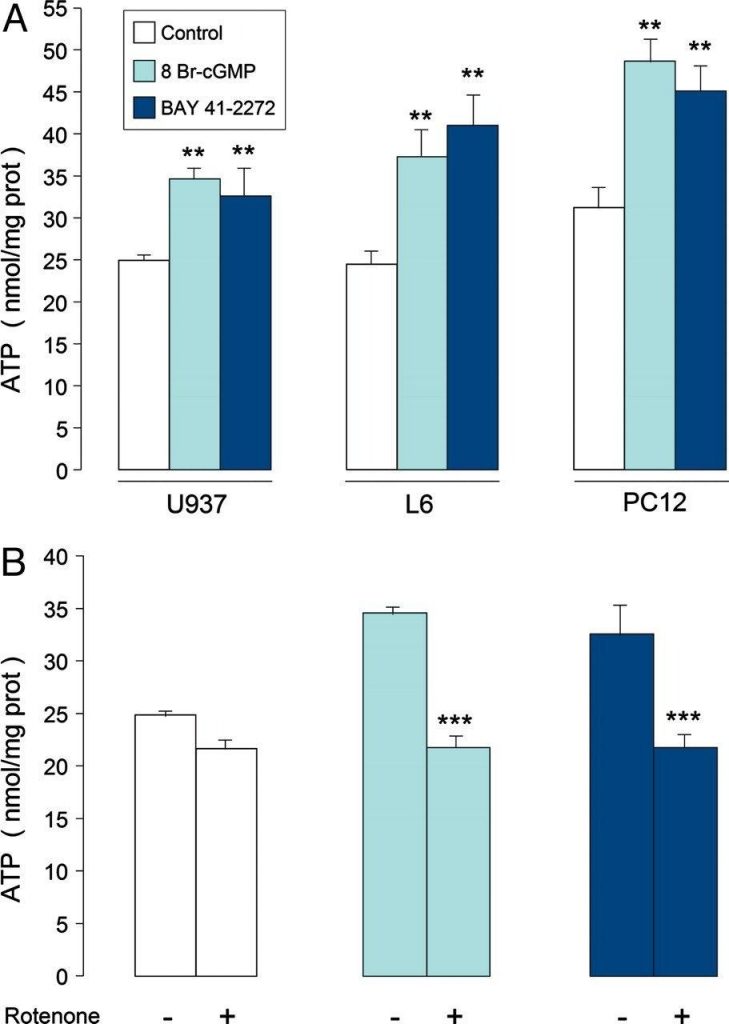
La biogénesis mitocondrial dependiente de cGMP conduce a aumentos en los niveles de ATP en estado estacionario a través de la fosforilación oxidativa. Las células U937, L6 y PC12 se cultivaron durante 6 días con o sin 3 mM de 8 Br-cGMP o 1 μM de BAY 41-2272, como se indica. (A) Niveles celulares de ATP en estado estacionario en las diversas preparaciones celulares. (B) Niveles de ATP en estado estacionario, medidos en células U937 en presencia o ausencia del inhibidor del complejo I rotenona (1 μM). Los asteriscos indican significación estadística, como se describe en Materiales y métodos, frente a controles (n = 3).
Las células inmortalizadas en cultivo normalmente cubren sus requerimientos energéticos a partir de la glucólisis, lo que representa la formación de ≈90% del ATP total ( 30 ). Por lo tanto, la exposición a corto plazo a un inhibidor de la cadena respiratoria debería proporcionar solo efectos menores en células en las que el acervo de ATP depende principalmente de la glucólisis, mientras que deberían ocurrir efectos mucho mayores en condiciones en las que la contribución del ATP generado mitocondrialmente se vuelve más significativa. Encontramos que la exposición de células U937 no tratadas al inhibidor del complejo I rotenona (1 μM durante 60 min) suprimió el consumo de O 2 (datos no mostrados) pero disminuyó el contenido celular de ATP solo marginalmente (en un 12,7%; ). Sin embargo, la rotenona causó una disminución mucho mayor en los niveles de ATP en células que habían sido tratadas durante 6 días con 8 Br-cGMP o BAY 41-2272, de modo que fueron los mismos que los detectados en células no tratadas. El tratamiento de cada tipo celular con desoxiglucosa (5 mM) durante 60 min produjo una marcada disminución del contenido de ATP, de modo que se mantuvieron cantidades similares en las células control y en las tratadas con 8 Br-cGMP o BAY 41-2272 (datos no mostrados). Estos resultados sugieren firmemente que el aumento del contenido de ATP mediado por cGMP puede atribuirse a la fosforilación oxidativa.
Para confirmar estas observaciones, se midieron las concentraciones de lactato en el medio extracelular/intracelular para obtener una estimación del flujo glucolítico, en condiciones idénticas a las utilizadas para la determinación de ATP. Las concentraciones de lactato fueron similares en las diferentes condiciones de tratamiento, tanto en el medio extracelular (valores de 3,82 ± 0,27 mM y 3,91 ± 0,25 mM en células no tratadas y en 8 células tratadas con Br-cGMP, respectivamente; n = 4) como dentro de las células (valores de 0,098 ± 0,015 μmol/mg de proteína y 0,099 ± 0,013 μmol/mg de proteína en células no tratadas y en 8 células tratadas con Br-cGMP, respectivamente; n = 4). Además, la actividad de dos enzimas clave de la glucólisis [a saber, hexoquinasa (0,104 ± 0,019 unidades/mg de proteína y 0,0985 ± 0,016 unidades/mg de proteína en células no tratadas y 8 células tratadas con Br-cGMP, respectivamente; n = 4) y gliceraldehído fosfato deshidrogenasa (1,005 ± 0,021 unidades/mg de proteína y 1,025 ± 0,14 unidades/mg de proteína en células no tratadas y 8 células tratadas con Br-cGMP, respectivamente; n = 4)] no mostró variaciones apreciables. Se obtuvieron resultados similares a los observados en células U937 en células L6 y PC12 (datos no mostrados). Por lo tanto, la biogénesis mitocondrial desencadenada por cGMP da como resultado mitocondrias funcionalmente activas, de modo que las células consumen más O2 y generan más ATP mediante un proceso independiente de la glucólisis.
Estudios previos en ratones eNOS –/– han indicado que el NO regula la biogénesis mitocondrial in vivo. Por lo tanto, hemos investigado la relación entre la biogénesis mitocondrial, el consumo de O2 y la generación de ATP en el cerebro, el corazón, el hígado, el riñón y el músculo gastrocnemio de ratones WT y eNOS –/– . El contenido mitocondrial basal (evaluado midiendo el ADNmt, las tasas de consumo basal de O2 y el contenido de ATP) se redujo en los tejidos de los ratones eNOS –/– en comparación con sus contrapartes WT (consulte la Tabla 2). Estos resultados indican que la generación de NO por eNOS in vivo es necesaria para mantener mitocondrias funcionalmente activas capaces de acoplar el aumento del consumo de O2 con la generación de ATP. El menor contenido mitocondrial en ratones eNOS –/– estuvo acompañado en todos los tejidos por menores niveles de ARNm de PGC-1α, compatibles con la participación de la expresión de PGC-1α en la biogénesis mitocondrial tanto en los tejidos como en las células (Tabla 2).
Por último, comparamos ratones WT y eNOS –/– mediante análisis morfométrico de mitocondrias a nivel ultraestructural. Los resultados obtenidos en dos tejidos representativos (hígado y gastrocnemio) se informan en la Tabla 1 , y las micrografías representativas se muestran en la Fig. En los hepatocitos de animales WT, las mitocondrias se empaquetaron uniformemente en todo el citoplasma y tenían forma redonda con disposición regular de crestas. En los hepatocitos de ratones eNOS –/–, las mitocondrias fueron significativamente más pequeñas y menos densamente empaquetadas. Dos grupos mitocondriales claramente distintos estaban presentes en las fibras musculares (uno intramiofibrilar y otro subsarcolemal). El análisis morfométrico del grupo intramiofibrilar dio resultados similares a los observados en el hígado, porque las mitocondrias de los tejidos eNOS –/– fueron significativamente más pequeñas y menos densamente empaquetadas. Este hallazgo estuvo acompañado de una marcada disminución en el número de mitocondrias residentes en la región subsarcolemal (Fig. 5).
Discusión
Nuestros resultados actuales muestran que la biogénesis mitocondrial, evaluada mediante mediciones de la expresión de mtADN, COX-IV y Cyt c, fue inducida de manera similar por DETA-NO en tres tipos celulares de diferentes especies y linajes (es decir, U937 monocítico humano, mioblastos L6 de rata y células neurosecretoras PC12). La biogénesis mitocondrial estuvo acompañada por una mayor expresión de PGC-1α, NRF-1 y Tfam (aunque en grados variables en los diferentes tipos celulares), y fue mediada por cGMP porque 8 Br-cGMP imitó, y el inhibidor de sGC ODQ abolió, los efectos de DETA-NO. La observación de que BAY 41-2272 desencadenó la biogénesis mitocondrial por medio de la misma vía de señalización activada por NO confirmó el papel clave de cGMP en este proceso. Se estableció además que el NO endógeno desempeña un papel crucial en la biogénesis mitocondrial en modelos animales, ya que el contenido mitocondrial de diversos tejidos de ratones eNOS –/– fue significativamente menor que el de los animales WT. Estos resultados, junto con los resultados, demuestran claramente que la biogénesis mitocondrial es un fenómeno general que ocurre en células en cultivo y tejidos animales, probablemente a través de la misma cascada de eventos de señalización.
Aunque los aumentos inducidos por NO/cGMP en el contenido de mtADN, Cyt c, COX-IV PGC-1α, Tfam y NRF-1 indican estimulación de la biogénesis mitocondrial, estos aumentos no implican necesariamente que las mitocondrias resultantes sean funcionales. Por lo tanto, medimos dos parámetros de la función mitocondrial (es decir, consumo de O2 y producción de ATP), y descubrimos que la biogénesis mitocondrial inducida por NO/cGMP estaba acompañada de un mayor consumo de O2 a través de la respiración celular acoplada funcionalmente vinculada a una mayor producción de ATP. Curiosamente, las líneas celulares utilizadas son altamente glucolíticas, pero aún responden a cGMP aumentando la fosforilación oxidativa. El hecho de que no observáramos una inhibición concomitante en la tasa de producción de ATP por glucólisis puede atribuirse al hecho de que dicho control de la glucólisis es bajo en las células derivadas de tumores altamente glucolíticas que utilizamos.
Se sabe que el cGMP regula diversas vías de señalización intracelular implicadas en procesos biológicos clave, como la proliferación y diferenciación celular. Por lo tanto, no sorprende que, además de la biogénesis mitocondrial, hayamos observado cambios inducidos por NO/cGMP en el contenido proteico celular o en las tasas de proliferación. Sin embargo, a diferencia de los cambios constantes en el consumo de O₂ y el contenido de ATP, los cambios en el contenido proteico celular o en las tasas de proliferación fueron variables y significativamente menores en los tres tipos celulares investigados. Estos resultados sugieren firmemente que los aumentos en el consumo de O₂ y el contenido de ATP no son simplemente consecuencia de cambios en el contenido proteico celular o en las tasas de proliferación.
La caracterización funcional de las mitocondrias en tejidos animales confirmó y amplió los resultados que obtuvimos con las células en cultivo. El contenido mitocondrial reducido en tejidos de ratones eNOS –/– estuvo acompañado de una reducción tanto del consumo basal de O 2 como de los niveles de ATP en estado estacionario, asociados con una reducción significativa en el tamaño mitocondrial. Estas observaciones ocurrieron en tejidos que dependen del metabolismo oxidativo (como el cerebro) y en tejidos glucolíticos (como el músculo gastrocnemio), lo que indica que el efecto del NO en la biogénesis mitocondrial es un fenómeno general. Curiosamente, en el músculo gastrocnemio, se ha demostrado que la biogénesis mitocondrial es importante en la conversión de fibras de tipo II (glucolíticas) en fibras de tipo I (oxidativas).
Existe un alto grado de flexibilidad en el tamaño, número y masa mitocondrial, que están mediados por una red compleja y estrechamente coordinada de diferentes vías reguladoras. Se ha demostrado que el volumen mitocondrial y la extensión de la red ( se correlacionan positivamente con la eficiencia de la función respiratoria mitocondrial ). Por lo tanto, es probable que la disminución del tamaño mitocondrial observada en los tejidos de ratones eNOS –/– sea un correlato morfológico de la función mitocondrial reducida. El grupo subsarcolemal del músculo gastrocnemio parece ser particularmente dinámico en la adaptación a los cambios crónicos a nivel de la actividad contráctil, y parece contener principalmente mitocondrias en una etapa más temprana de la biogénesis. Estos hallazgos se correlacionan bien con nuestra observación de que este grupo de mitocondrias se ve más afectado por la privación de NO que las mitocondrias intramiofibrilar.
Nuestros resultados obtenidos de ratones eNOS –/– en condiciones de reposo demuestran que la generación endógena de NO por eNOS es necesaria para el mantenimiento in vivo del contenido mitocondrial basal. Esta consideración no descarta la posibilidad de que otras isoformas de NOS puedan participar en la biogénesis mitocondrial cuando este proceso necesita intensificarse. Por ejemplo, la activación de NOS neuronal que ocurre durante el ejercicio del músculo esquelético podría contribuir a la biogénesis mitocondrial dependiente de PGC-1α inducida por el ejercicio. Tampoco podemos descartar por completo la posibilidad de que la reducción del flujo sanguíneo secundario a la deficiencia de eNOS contribuya indirectamente al fenotipo observado. Sin embargo, esta eventualidad es poco probable porque no se observaron signos importantes de atrofia en los tejidos investigados.
Nuestros resultados pueden tener consecuencias importantes en términos de biología celular y tisular, porque la actividad mitocondrial y la biogénesis desempeñan un papel crítico en varios procesos, incluyendo la adquisición de características de células de grasa parda por adipocitos blancos, el cambio de fibras musculares esqueléticas de metabolismo glucolítico a oxidativo, y la regeneración de músculos cardíacos y esqueléticos. Además, el deterioro de la función mitocondrial está asociado con enfermedades neurodegenerativas, trastornos neuromusculares, insuficiencia hepática y cardíaca, y diabetes tipo 2. Por lo tanto, la posibilidad de generar nuevas mitocondrias metabólicamente activas podría mejorar el resultado de estas patologías. En este contexto, es interesante que BAY 41-2272 y los nuevos compuestos donantes de NO tengan una posible aplicación clínica.
En conclusión, nuestros resultados, combinados con evidencia previa, indican que el NO regula el metabolismo celular mediante dos acciones distintas a nivel mitocondrial. Por lo tanto, la detección aguda de O₂ por parte de las células, regulada por la unión reversible del NO a la COX-IV con la consiguiente reducción del consumo de O₂ ( 6 , 10 , 11 ) , parece complementarse con cambios a largo plazo que dependen de la biogénesis mitocondrial. Queda por investigar cómo estas dos acciones pueden coordinarse para satisfacer las demandas energéticas de la célula en diferentes condiciones.
Abreviaturas: sGC, guanilato ciclasa “soluble”; PGC-1α, coactivador 1α del receptor activado por el proliferador de peroxisomas γ; NRF-1, factor respiratorio nuclear 1; Tfam, factor de transcripción mitocondrial A; eNOS, NO sintasa endotelial; DETA-NO, (Z)-1-[2-(2-aminoetil)- N -(2-amonioetil) amino]diazen-1-io-1,2 diolato; ODQ, H -(1,2,4)-oxadiazolo [4,3-]quinoxalin-1-ona; Cyt c, citocromo c; FCCP, cianuro de carbonilo p -trifluorometoxi fenilhidrazona.
Bibliografía de referencia
- Kann, O. y Kovács, R. Mitocondrias y actividad neuronal. Am. J. Physiol. Cell Physiol. 292, C641-57 (2007).
- Yu-Wai-Man, P., Griffiths, PG y Chinnery, PF. Neuropatías ópticas mitocondriales: mecanismos patológicos y estrategias terapéuticas. Prog. Retin. Eye Research 30 , 81–114 (2011).
- Reeve, AK et al. Disfunción mitocondrial en las sinapsis de neuronas de la sustancia negra en la enfermedad de Parkinson. NPJ Parkinsons Dis. 4, 9 (2018)
- Das, A. e Imanishi, Y. Estrategias de descubrimiento de fármacos para degeneraciones retinianas hereditarias. Biology 11, 1338 (2022)
- Neishabouri, A. y Faisal, AA. Conducción saltatoria en axones amielínicos: la agrupación de canales de Na+ en las balsas lipídicas permite la conducción microsaltatoria en las fibras C. Front. Neuroanat. 8, 109 (2014)
- García, D. y Shaw, RJ AMPK: mecanismos de detección de energía celular y restauración del equilibrio metabólico. Mol. Cell 66, 789–800 (2017)
- Zhao, P. et al. TBK1 en la encrucijada de la inflamación y la homeostasis energética en el tejido adiposo. Cell 172 , 731–743.e12 (2018).
- Mattson, MP y Arumugam, TV. Características del envejecimiento cerebral: modificación adaptativa y patológica por estados metabólicos. Cell Metab. 27 , 1176–1199 (2018)
- Sluch, VM et al. Mejora de la diferenciación de células madre e inmunopurificación de células ganglionares de la retina humana modificadas genéticamente. Stem Cells Transl. Med. 6, 1972-1986 (2017)
- Zhang, KY et al. Función de la membrana limitante interna en el injerto estructural y el espaciamiento topográfico de células ganglionares de la retina derivadas de células madre humanas trasplantadas. Stem Cell Rep. 16, 149–167 (2021)
- Das, A., Bell, CM, Berlinicke, CA, Marsh-Armstrong, N. y Zack, DJ. El cambio programado en las vías de degradación mitocondrial durante la diferenciación de las células ganglionares de la retina humana a partir de células madre es crucial para la supervivencia de las CGR. Redox Biol. 34, 101465 (2020).
- VanderWall, KB et al. Las células ganglionares de la retina con una mutación en el gen OPTN(E50K) asociado al glaucoma presentan fenotipos neurodegenerativos al derivar de organoides retinianos tridimensionales. Stem Cell Rep. 15, 52–66 (2020)
- Gureev, AP, Shaforostova, EA y Popov, VN. Regulación de la biogénesis mitocondrial como vía para la longevidad activa: Interacción entre las vías de señalización Nrf2 y PGC-1α. Portada. Genet. https://doi.org/10.3389/fgene.2019.00435 (2019).

Alfonso Barbosa Domínguez. Médico y Cirujano – Universidad del Valle, Cali Colombia. Ingeniero Químico y Tecnólogo en Alimentos – Universidad del Valle, Cali Colombia. Profesional en Deporte y Actividad Física – Escuela Nacional del Deporte, Cali Colombia. Tecnólogo Deportivo con énfasis en Deportes Acuáticos. Escuela Nacional del Deporte- Cali Colombia.
Especializaciones:
Biotipología y Antropometría Aplicada al Deporte (Brasil),
Ciencias Biomédicas Aplicadas al Deporte. Universidad del Cauca Popayán Colombia.
Maestría en:
Fisiología Aplicada al Deporte – Universidad de Kiev, Ucrania.
Doctorado en:
Neurofisiología Aplicada al Deporte – Universidad de Colonia, Alemania.
Postdoctorado en:
Epigenética – Universidad Leipzig, Alemania.
Ex campeón suramericano y centroamericano de natación.
Nadador master del club Aquaticgym

